Recent evolution of the ability to read and write
Recent evolution of the ability to read and write
The Visual Word Form Area provides an interesting case of gene-culture coevolution.

Written by Peter Frost.
The Visual Word Form Area is a brain region that helps us recognize written words and letters. (We can read without it, albeit less easily.) When a man suffered an accidental lesion to his VWFA during brain surgery, he lost much of his reading ability but experienced no loss of object recognition and naming, face processing or general language abilities. Six months later he had partially recovered, but reading still took twice as long as it had before the surgery (Gaillard et al, 2006).
The VWFA is composed of neurons that were once used for face recognition:
Thus, learning to read must involve a ‘neuronal recycling’ process whereby pre-existing cortical systems are harnessed for the novel task of recognizing written words. … [Such areas of the cortex] possess the appropriate receptive fields to recognize the small contrasted shapes that are used as characters, and the appropriate connections to send this information to temporal lobe language areas (Dehaene & Cohen, 2011)
This “recycling” seems to have become hardwired, at least in some people. In an MRI study, the VWFAs of Swiss preschoolers preferentially responded to pictures of letter strings once the children had played a grapheme/phoneme correspondence game for a total of 3.6 hours over an 8-week period. Yet only a few of them could actually read and even then only at a rudimentary level (Brem et al., 2010).
It may therefore be that humans first identified words by using face-recognition neurons. Then, as the ability to read became more important, natural selection favored those individuals who could free up more of those neurons for reading. The eventual outcome was a large neuronal population dedicated solely to word recognition, i.e., the VWFA.
This evolutionary process may be broken down into four stages:
Reading without a VWFA. This took twice as long as reading with a VWFA, as shown in the case study above. Reading was particularly arduous in early times because texts were continuous streams of characters with little or no punctuation.
Need for people who could read and write. With the rise of the state and the development of trade, demand grew for people who could read and write large volumes of text without getting tired. Known as ‘scribes,’ they had to record transactions, decisions, and edicts for government and business. Many became further specialized as stenographers, copyists, secretaries, notaries, calligraphers and so on.
Expansion of the scribal class. Demand for scribes was driven by two positive feedback loops. First, as the number of texts grew, so did the number of old or damaged ones to be copied and updated. Second, as scribes themselves grew in number, they became a significant source of supply and demand. In addition to creating texts for their patrons, they became producers and consumers in their own right. Reading and writing went beyond record-keeping: it developed into an intellectual activity oriented toward fiction, history and philosophy.
Spread of reading and writing through the general population. The economic success of scribes enhanced their reproductive success, which led to “surplus” offspring who would enter the general population. Their genetic propensities thus increased in frequency within the entire gene pool, enabling more people to read and write with less effort. The culture itself began to change. Because information was now being accumulated on “hard copy,” people could draw on a larger store of knowledge. They could learn not only from the living but also from the dead.

The above evolutionary model may seem hard to believe. How could natural selection create a new mental organ in such a short time? “The invention of writing is too recent and, until the last century, concerned too small a fraction of humanity to have influenced the human genome” (Dehaene & Cohen, 2011). Writing emerged in the Middle East about six thousand years ago, and some societies adopted writing only within the past century. Even in societies with a long literary tradition, reading and writing were confined to a minority until recent times.
But such rapid evolution is not as paradoxical as it may seem. First, the VWFA did not evolve from scratch. A new task had to be performed by an existing population of neurons, which at first did it clumsily but then evolved to do it better. This sort of repurposing has been common in our evolutionary history (Harpending & Cochran, 2002).
Second, selection can change an entire population even if it acts only within a small subgroup: so long as that subgroup is more reproductively successful than the rest of the population, its genetic characteristics will become more prevalent. This has been shown by Gregory Clark’s work on recent evolution in English population. As late as the 19th century, the lower class did not replace itself demographically and was continually replenished by downwardly mobile individuals from the middle class. Though initially small, the middle class grew through a higher rate of natural increase until its lineages dominated the English gene pool. Thus, the entire population became shaped by selection pressures that were confined to an initially small subgroup (Clark, 2007). In the ancient world, scribes similarly enjoyed elevated reproductive success, as evidenced by the Book of Sirach: “If [a scribe] lives long, he will leave a name greater than a thousand.” In other words, his descendants will be numerous.
Third, although only a minority were fully literate, many more were partially literate. The literacy rate of past societies is estimated from signed documents of one sort or another: wills, court depositions, marriage certificates and so on (Barr & Kamil, 1996, p. 52). If the signatory wrote an ‘X’, he or she is deemed to have been illiterate. Although such people undoubtedly had trouble writing, especially writing in cursive script, they may have been able to read texts of block letters, such as those on storefronts. This view is supported by the discovery of ancient graffiti, such as that at Pompeii, which seem to be written by ordinary people, including prostitutes. Current historical methods thus underestimate the number of people with at least some reading and writing ability.
In this, and many other ways, humans have directed their evolution. First they perform a new task by leveraging their phenotypic plasticity. This new way of doing things then acts as a template for natural selection. Over generations, the new phenotype gradually becomes a new genotype.
Does the ability to read and write vary within our species?
If humans evolved to become better at reading and writing, this may have happened to different extents in different populations. In particular, the VWFA should be larger and more specialized in those populations that have been literate or partially literate for longer.
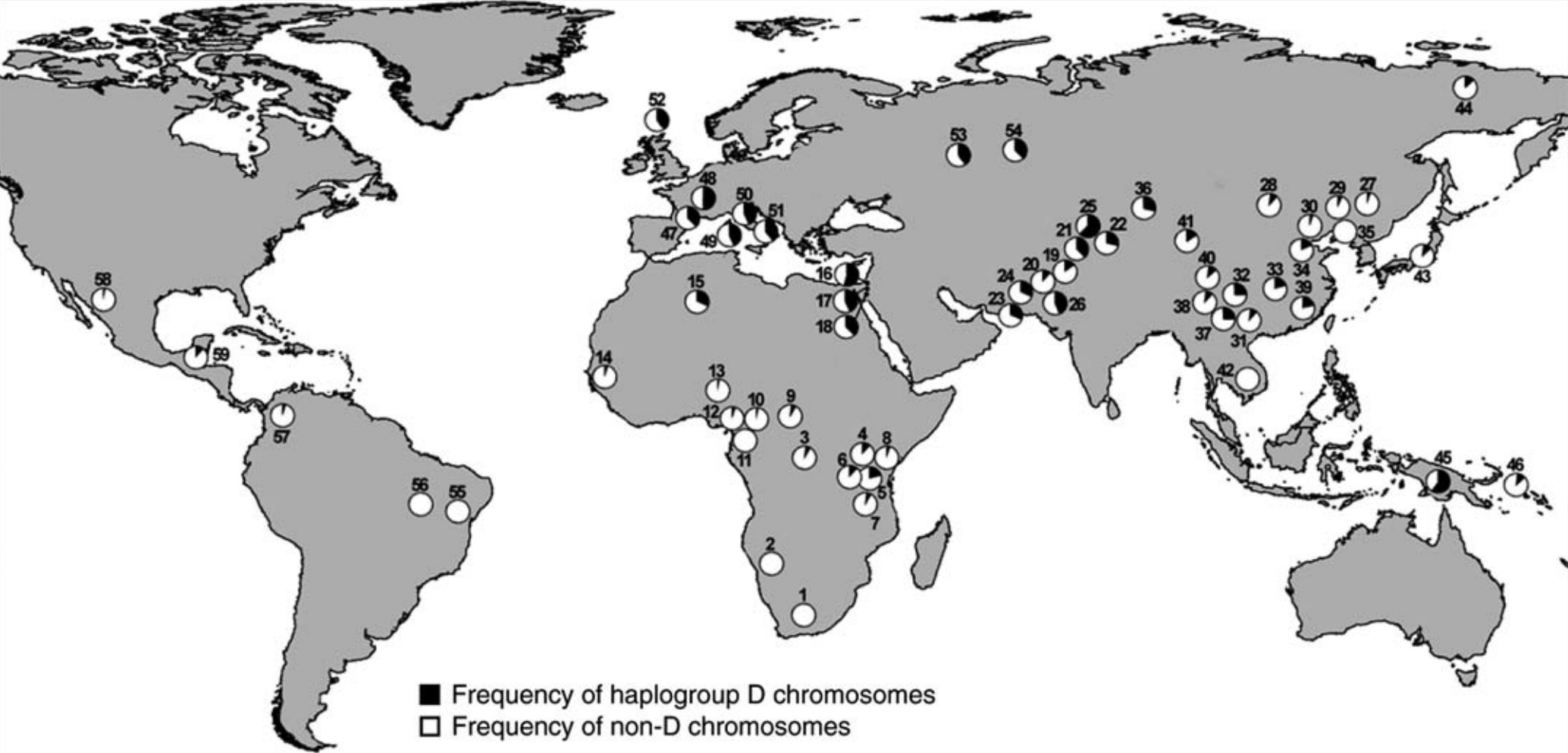
Differential selection for reading ability is indicated by two lines of evidence: 1) the geographic distribution of the latest ASPM variant; and 2) the differences among populations in the degree of competition between face recognition and word recognition.
ASPM is a gene that helps regulate primate brain growth (Wu et al., 2023). In humans, a new variant arose some six thousand years ago in the Middle East, where it is still most prevalent (37-52%). It then spread outward – more so into Europe (38-50%) than into East Asia (0-25%) (Mekel-Bobrov et al., 2005). This variant thus seems to track the spread of writing within Eurasia, particularly the spread of alphabetical script from an origin in Mesopotamia c. 5,400 to 5,100 years ago (Frost, 2011).
When this geographic pattern was discovered two decades ago, it was thought to be concrete evidence of recent cognitive evolution. Nonetheless, two research teams failed to find any association between the latest ASPM variant and cognitive ability (Mekel-Bobrov et al. 2007; Rushton et al. 2007). Philippe Rushton argued against pursuing further research, on the assumption that IQ includes all aspects of mental effort. In an email he told me: "Generally there isn't thought to be much left to be explained after g is taken out."
Researchers have since found that cognitive ability (g) does not correlate with face recognition; the two seem to function independently of each other (Zhu et al., 2010). The same should be true for cognitive ability and word recognition, since we recognize words by using neurons that evolved out of face-recognition neurons.
So the question is still open. We should pursue this abandoned line of research to see whether the presence or absence of the latest ASPM variant correlates with performance on word recognition, particularly words based on sound-based characters. Indeed, the geographic pattern seems to mirror the historical spread of alphabetical script, which puts much more demand on the VWFA than does ideographic script. Whereas Westerners process their sound-based characters only in the VWFA, the Chinese process their idea-based characters with assistance from other brain regions (Liu et al., 2008).
If the VWFA is a product of recent human evolution, it may be more developed in some populations than in others. The more your VWFA has had time to evolve, the more you will recognize words without loss of face recognition. The less time it has had to evolve, the more these two tasks will compete against each other for access to the same limited number of neurons.
Thus, in populations where many people began to read and write only in recent times, illiterates should suffer some loss of face recognition if they learn to read. Conversely, they should suffer no such loss in a population that has long had reading and writing.
Both types of population have provided participants for MRI studies on the degree of competition between word recognition and face recognition.
First, let’s consider populations with a short history of reading and writing. In four studies, learning to read incurred some loss of face recognition. The participants were wholly or mostly Brazilian:
Dehaene et al., 2010 – Portugal/Brazil, illiterate or ex-illiterate adults, n=63. Among unschooled adults, ex-illiterates had more trouble than illiterates in recognizing faces. Word recognition seemed to compete with face recognition on the periphery of the VWFA.
Pegado et al., 2014 – Portugal/Brazil, 49 illiterates and ex-illiterates, n=49 (mostly from the previous study). Literacy seemed to “push” face recognition away from the VWFA and toward the right hemisphere.
Braga et al., 2017 – Brazil, illiterate man who had never gone to school, n=1. Faces and words seemed to compete for processing within the VWFA. The authors concluded “that faces and written words occupy distinct but neighboring locations in the left occipitotemporal cortex, and that the growth of word-related responses induces a shift in the boundary between these two preferences and thus a relative decrease in the left-hemispheric response to faces.”
de Andrade & França, 2020 – Brazil (Rio de Janeiro), children, 4 to 8 years old, n=30. The participants lost some ability to recognize new faces when learning to read and write. This learning task helped them process speech, but adversely affected their capacity to recognize faces that had undergone alterations to the eyes, nose, and mouth. This adverse impact was still present among the oldest students in the second grade.
Now let’s consider populations with a long history of reading and writing. In two studies, learning to read incurred no loss of face recognition. The participants were either South Asian or Danish.
Hervais-Adelman et al., 2019 – India (near Lucknow), literate and illiterate adults, n=91. The authors concluded “that far from cannibalizing the territory of its neighbors, the VWFA is rather overlaid upon these and not only remains responsive to other categories of visual stimuli but also recruits an assembly of neural populations in much the same way, regardless of whether the input is orthographic, albeit with a lesser overall response magnitude.”
Kühn et al., 2021 – Denmark (Copenhagen), children 5-7 years old, n=82. The authors found no loss of face recognition when they considered all of the schoolchildren learning to read, although a quarter of them did show some loss.
In summary, the VWFA seems to be larger and more specialized in populations with a longer history of full or partial literacy, such as Danes and South Asians. Conversely, it seems to be smaller and less specialized in populations with a shorter history of full or partial literacy, such as Brazilians – the majority of whom are at least partly African or Amerindian by descent.
The VWFA also seems to vary within populations. Although Danes have a relatively long history of full or partial literacy, about a quarter of the Danish schoolchildren had some VWFA inadequacy. This subgroup was likely of Danish descent according to a personal communication from one of the study’s authors.
Further evidence of population differences comes from a study of New York City elementary schoolchildren, which found differences in VWFA activation that correlated with socioeconomic status (SES). In students from high SES families, activation seemed to be more hardwired and less dependent on familiarity with the way sounds are visually represented:
In these [lower SES] children, phonological skill level is positively predictive of activation in the left fusiform region that supports rapid visual word recognition. … In contrast, as the SES of the population increases, children demonstrating a similar range of phonological skill show an attenuated brain–behavior relationship in this region. (Noble et al., 2006).
The study’s authors reported the ethnic breakdown of their sample: approximately one third Euro-American, one third African American, and one third other groups. It would have thus been possible to see whether ethnicity was better than SES at explaining the activation differences. Unfortunately, this was not done.
Parting thoughts
Much confusion is still caused by the assumption that the ability to recognize words cannot be hardwired, since reading and writing are too recent to have played any role in human evolution. This assumption is due partly to a broader assumption that human evolution essentially ended in the Paleolithic, and partly to the troubling inference that some humans, and some human populations, are inherently less capable of reading and writing. So we continue with a model where all children, everywhere, and at an unspecified early stage of development, repurpose some of their face-recognition neurons to create their own VWFA. This model is untenable, yet the obvious alternative is not considered, as a recent review article indicates (Rossion & Lochy, 2022).
To test the alternative model, we should first find out whether the ability to recognize words is associated with the latest ASPM variant. If an association is found, this model could then be applied to another region of the human brain, Exner’s area, which seems to control handwriting: if it is damaged, one loses the ability to write words and numbers by hand, while still being able to type them on a computer keyboard (Klein et al., 2016).
With Exner’s area, we again have an apparent case of recent evolution, specifically one where humans have coevolved with their cultural environment during the course of recorded history.
Peter Frost has a PhD in anthropology from Université Laval. His main research interest is the role of sexual selection in shaping highly visible human traits, notably skin color, hair color and eye color. Other research interests include gene-culture coevolution. Find his newsletter here.
Consider supporting Aporia with a paid subscription:

You can also follow us on Twitter.
References
Barr, R. & Kamil, M.L. (1996). Handbook of Reading Research vol. 2, Routledge.
Braga, L. W., Amemiya, E., Tauil, A., Suguieda, D., Lacerda, C., Klein, E., ... & Dehaene, S. (2017). Tracking adult literacy acquisition with functional MRI: A single-case study. Mind, Brain, and Education, 11(3), 121-132. https://doi.org/10.1111/mbe.12143
Brem, S., Bach, S., Kucian, K., Guttorm, T.K., Martin, E., Lyytinen, H., Brandeis, D., & Richardson, U. (2010). Brain sensitivity to print emerges when children learn letter-speech sound correspondences. Proceedings of the National Academy of Sciences U.S.A., 107(17), 7939–7944. https://doi.org/10.1073/pnas.0904402107
Clark, G. (2007). A Farewell to Alms. A Brief Economic History of the World, Princeton University Press, Princeton and Oxford. https://press.princeton.edu/books/paperback/9780691141282/a-farewell-to-alms
de Andrade, I. R., & França, A. I. (2020). A influência da alfabetização nas cognições de reconhecimento de faces e dos sinais da fala: um estudo psicolinguístico. The influence of literacy on face recognition and speech sign cognitions: a psycholinguistic study. Revista Prolíngua–ISSN, 1983(9979), 101.
Dehaene, S. & Cohen, L. (2011). The unique role of the visual word form area in reading, Trends in Cognitive Sciences, 15(6), 254-262. https://doi.org/10.1016/j.tics.2011.04.003
Dehaene, S., Pegado, F., Braga, L. W., Ventura, P., Filho, G. N., Jobert, A., ... & Cohen, L. (2010). How learning to read changes the cortical networks for vision and language. Science, 330(6009), 1359-1364. https://doi.org/10.1126/science.1194140
Frost, P. (2011). Human nature or human natures? Futures, 43, 740-748. http://dx.doi.org/10.1016/j.futures.2011.05.017
Gaillard, R., Naccache, L., Pinel, P., Clémenceau, S., Volle, E., Hasboun, D., ... & Cohen, L. (2006). Direct intracranial, FMRI, and lesion evidence for the causal role of left inferotemporal cortex in reading. Neuron, 50(2), 191-204. https://doi.org/10.1016/j.neuron.2006.03.031
Harpending, H., & Cochran, G. (2002). In our genes. Proceedings of the National Academy of Sciences USA, 99(1), 10-12. https://doi.org/10.1073/pnas.012612799
Hervais-Adelman, A., Kumar, U., Mishra, R. K., Tripathi, V. N., Guleria, A., Singh, J. P., ... & Huettig, F. (2019). Learning to read recycles visual cortical networks without destruction. Science Advances, 5(9). https://doi.org/10.1126/sciadv.aax0262
Klein, E., Willmes, K., Jung, S., Huber, S., Braga, L. W., & Moeller, K. (2016). Differing connectivity of Exner’s area for numbers and letters. Frontiers in Human Neuroscience, 10, 281. https://doi.org/10.3389/fnhum.2016.00281
Kühn, C. D., Wilms, I. L., Dalrymple, K. A., Gerlach, C., & Starrfelt, R. (2021). Face recognition in beginning readers: Investigating the potential relationship between reading and face recognition during the first year of school. Visual Cognition, 29(4), 213-224. https://doi.org/10.1080/13506285.2021.1884151
Liu, C., Zhang, W. T., Tang, Y. Y., Mai, X. Q., Chen, H. C., Tardif, T., & Luo, Y. J. (2008). The visual word form area: evidence from an fMRI study of implicit processing of Chinese characters. Neuroimage, 40(3), 1350-1361. https://doi.org/10.1016/j.neuroimage.2007.10.014
Mekel-Bobrov, N., Gilbert, S. L., Evans, P. D., Vallender, E. J., Anderson, J. R., Hudson, R. R., ... & Lahn, B. T. (2005). Ongoing adaptive evolution of ASPM, a brain size determinant in Homo sapiens. Science, 309(5741), 1720-1722. https://doi.org/10.1126/science.1116815
Mekel-Bobrov, N., Posthuma, D., Gilbert, S. L., Lind, P., Gosso, M. F., Luciano, M., ... & Lahn, B. T. (2007). The ongoing adaptive evolution of ASPM and Microcephalin is not explained by increased intelligence. Human molecular genetics, 16(6), 600-608. https://doi.org/10.1093/hmg/ddl487
Noble, K. G., Wolmetz, M. E., Ochs, L. G., Farah, M. J., & McCandliss, B. D. (2006). Brain–behavior relationships in reading acquisition are modulated by socioeconomic factors. Developmental Science, 9(6), 642-654. https://doi.org/10.1111/j.1467-7687.2006.00542.x
Pegado, F., Comerlato, E., Ventura, F., Jobert, A., Nakamura, K., Buiatti, M., ... & Dehaene, S. (2014). Timing the impact of literacy on visual processing. Proceedings of the National Academy of Sciences, 111(49), E5233-E5242. https://doi.org/10.1073/pnas.1417347111
Rossion, B., & Lochy, A. (2022). Is human face recognition lateralized to the right hemisphere due to neural competition with left-lateralized visual word recognition? A critical review. Brain Structure and Function, 227(2), 599-629. https://doi.org/10.1007/s00429-021-02370-0
Rushton, J. P., Vernon, P. A., & Bons, T. A. (2007). No evidence that polymorphisms of brain regulator genes Microcephalin and ASPM are associated with general mental ability, head circumference or altruism. Biology Letters, 3(2), 157-160. https://doi.org/10.1098/rsbl.2006.0586
Wu, X., Li, Z., Wang, Z. Q., & Xu, X. (2023). The neurological and non-neurological roles of the primary microcephaly-associated protein ASPM. Frontiers in Neuroscience, 17, 1242448. https://doi.org/10.3389/fnins.2023.1242448
Zhu, Q., Song, Y., Hu, S., Li, X., Tian, M., Zhen, Z., ... & Liu, J. (2010). Heritability of the specific cognitive ability of face perception. Current Biology, 20(2), 137-142. https://doi.org/10.1016/j.cub.2009.11.067
Source: Aporia


{kind=link}
Comments
Post a Comment